VEDIAMO ADESSO, CON QUESTO SPECIFICO ARTICOLO DEL Dott.
Francesco Pellegrino, COME AVVIENE NEL DETTAGLIO
LA CONTRAZIONE MUSCOLARE
, “CHE COSA” E “COME”
LA SI DETERMINA
; MA SOPRATUTTO “ PERCHE’ ”… UN VALIDO COMPENDIO COGNITIVO PER OGNI
ISTRUTTORE E PREPARATORE ATLETICO CHE SI POSSA DEFINIRE TALE.
LA CONTRAZIONE MUSCOLARE
Di: Dott. Francesco Pellegrino
Come avviene la contrazione muscolare
A questo punto,
dopo aver letto l’articolo precedente (La
muscolatura del corpo umano) saprete già che la contrazione della
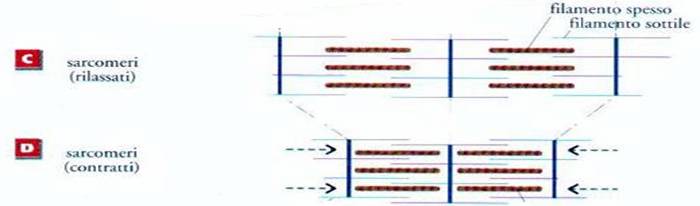
fibra muscolare è dovuta ad una diminuzione di lunghezza di ogni singolo
sarcomero, con accostamento di due bande A successive, per riduzione di
lunghezza del disco I. Sappiamo inoltre che la riduzione di I é determinata
dallo slittamento dei filamenti sottili nell’intervallo tra i filamenti
spessi.
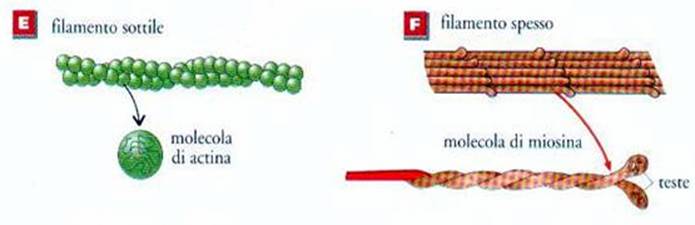
Ma si andrà
ancora di più nel dettaglio facendo osservare come le espansioni delle molecole
di miosina nel corso della contrazione si connettono con i miofilamenti sottili
di actina, tale connessione é data dal susseguirsi rapidissimo di legami
transitori tra actina e miosina; a questi legami si accompagnano rapide
oscillazioni delle espansioni dei filamenti di miosina che, quasi fossero
ciglia, fanno slittare il miofilamento sottile nell’intervallo tra i
miofilamenti spessi.
Studieremo
adesso più a fondo, i legami che si formano tra le “teste” delle molecole
di miosina e le molecole di actina. Intanto possiamo dire che questi legami,
temporanei, vengono detti “ponti trasversali actomiosinici”. Dobbiamo però
mettere l’accento… sul fatto che le espansioni globulari delle molecole di
miosina, oltre ad essere la sede dei legami con le molecole di actina durante la
contrazione, svolgono anche un’azione enzimatica, scindendo l’ATP in ADP,
reazione che fornisce l’energia necessaria alla contrazione stessa… e che
inoltre… è il legame di una nuova molecola di ATP con la molecola di miosina,
che ne libera l’estremità dal legame con la molecola di actina e la rende
perciò disponibile al legame successivo.
Per analizzare
“in dettaglio progressivo” come avvengono questa serie di reazioni… ci
avvarremo della proiezione della Fig. 6 (gia presentata anche nell’altro
articolo) di cui faremo adesso, la descrizione punto per punto:
Fig. 6
Schema del meccanismo con cui viene generata la forza nella contrazione
muscolare
-
partiamo dal
momento in cui una molecola di ATP si lega con la testa di una molecola di
miosina determinando la dissociazione del ponte trasversale tra questa e l’unità
del filamento di actina.
-
L’idrolisi
dell’ATP ad ADP + Pi fa si che l’asse della testa si sollevi rispetto all’asse
del filamento spesso, il muscolo è ancora a riposo.
-
Ora anche l’actina
è pronta (vedremo subito dopo in che senso) su di essa si instaura il legame
con il complesso testa della miosina - ADP - Pi.
-
L’interazione
actina - miosina causa il rilascio di Pi e dell’ADP dalla testa della miosina,
questo causa un ulteriore aumento dell’inclinazione dell’asse della testa
della miosina, che trascina con se nello spostamento, il filamento di actina a
cui è attaccata: è la fase in cui si genera la forza del muscolo.
-
Ora la testa
della miosina è di nuovo pronta a legarsi con una nuova molecola di ATP per
dare così il via ad un nuovo ciclo di contrazione.
Avevamo
lasciato in sospeso al punto 3, le modalità con cui l’actina si prepara ad
accogliere il legame con la testa della miosina. Per la trattazione di questo
argomento ci sarà utile la proiezione della Fig. 7.
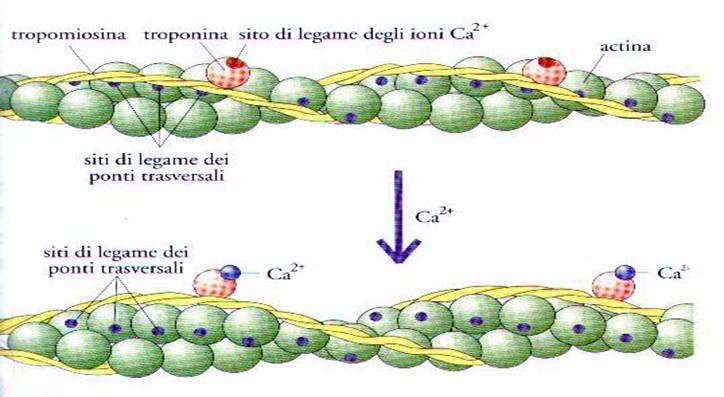
Fig. 7
Filamento di actina con le posizioni relative delle molecole di tropomiosina e
troponina prima e dopo il legame di quest’ultime con gli ioni Ca2+
Con l’aiuto
di questa figura si potrà comprendere come la contrazione muscolare sia
controllata da ioni Ca2+, contenuti nei vacuoli del reticolo che circonda le
miofibrille, e da due gruppi di molecole organiche:
-
la tropomiosina;
-
la troponina.
Dalla figura
inoltre, si nota anche la differente struttura delle 2 molecole: Tropomiosina:
che sono filamentose, rigide e costituite da due subunità avvolte ad elica l’una
sull’altra, che corrono lungo i filamenti di actina. Troponina: che sono
costituite da degli aggregati globulari proteici, legate sia alle molecole della
tropomiosina che a quelle dell’ actina e sono disposte ad intervalli regolari.
Nella parte superiore della Fig.
7 è illustrata la disposizione delle diverse
molecole quando
il muscolo è a riposo e si vede che i filamenti di tropomiosina sono disposti
lungo i filamenti di actina, in maniera tale da schermare i siti in cui si
stabilisce la connessione dei ponti trasversali e gli ioni Ca2+ non sono ancora
presenti, perché confinati all’interno dei vacuoli del reticolo
sarcoplasmatico. Perché avvenga una variazione in questa distribuzione,
occorrerà che intervenga un impulso “elettrico” proveniente dal sistema
nervoso. Nel momento in cui arriva alle miofibrille un potenziale d’azione
trasmesso dai neuroni, la permeabilità della membrana del reticolo
sarcoplasmatico agli ioni Ca2+ varia e permette che questi vengano rilasciati
dai piccoli serbatoi in cui sono contenuti, andando a stabilire dei legami con
le molecole di troponina.
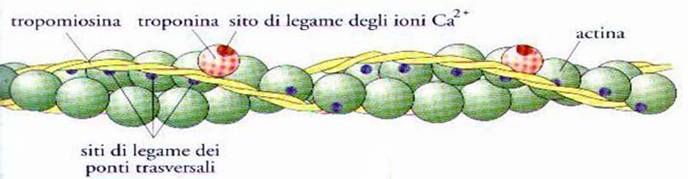
Gli effetti di
questo evento sono illustrati nella parte inferiore della Fig. 7.
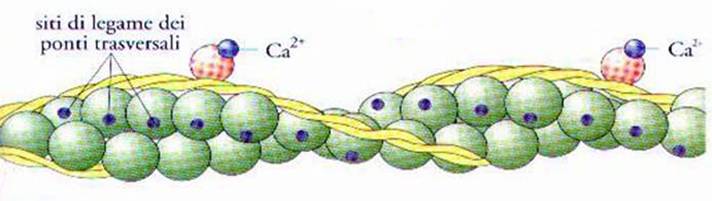
Si vede bene
infatti, come la costituzione di questo legame, fa si che muti la posizione
delle molecole di troponina ed essendo queste legate alle molecole di
tropomiosina e di actina, nel muoversi causano uno slittamento tra le due catene
molecolari. Lo slittamento delle molecole di tropomiosina relativamente a quelle
di actina, avviene in modo tale da scoprire i siti dei ponti trasversali, posti
sulle molecole di actina e queste sono così pronte per stabilire il legame con
le molecole di miosina. Questo avviene in modo da far si che l’actina sia
pronta al legame con la miosina ed è proprio questo, che si intendeva dire
nella fase 3 del ciclo visto sopra.
In conclusione
si dirà che: quando lo stimolo cessa e la permeabilità della membrana del
reticolo si riabbassa, esiste un sistema di pompaggio che riporta gli ioni Ca2+
all’interno dei serbatoi. La concentrazione di Ca2+ lungo i filamenti quindi
si riabbassa, la troponina si disattiva… e la contrazione cessa.
Terminazioni nervose per il controllo dell’attività muscolare
Saprete
sicuramente, che la trasmissione degli impulsi nervosi… avviene per la
propagazione del potenziale d’azione attraverso gli “assoni” dei neuroni
motori. Saprete anche che l’assone del neurone motorio è rivestito da una “guaina
mielinica”, quello che dobbiamo invece trattare in questa sede… è “cosa
succede” quando il neurone raggiunge i muscoli. A questo scopo faremo
riferimento alla Fig. 8 la quale mostra, con dettaglio crescente, come l’assone
in prossimità della sua terminazione, si ramifica e nella parte finale
fuoriesce dalla guaina mielinica. Si fa notare che l’espansione terminale del
neurone motore a questo punto, si inserisce a ridosso di una ristretta “zona
specializzata” della membrana plasmatica della fibra muscolare striata e che
viene detta “placca motrice”. C’è da aggiungere, che un muscolo ha sempre
molte più fibre muscolari di quante fibre nervose motorie riceve; perciò…
una fibra nervosa forma più placche motrici e controlla più fibre muscolari.
Chiameremo “unità motoria” l’insieme di un neurone motorio e delle fibre
muscolari che esso innerva. Le unità motorie non hanno la stessa dimensione in
tutti i muscoli dell’organismo; in alcuni muscoli sono costituite da molte
fibre muscolari, fino a 1700; in altri da pochissime fibre muscolari, 6 o 7
soltanto. A un più elevato numero di unità motorie, in relazione al numero
totale di fibre muscolari, corrisponde una migliore modulazione della capacitá
contrattile del muscolo.
Lo stimolo
proveniente da un neurone motorio, determina la contrazione di tutte le fibre
muscolari ad esso collegate. Mentre la contrazione di un muscolo, a seconda
della sua entità o della forza che deve sviluppare, interessa un numero
variabile di unità motorie.
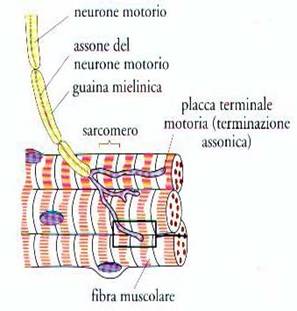
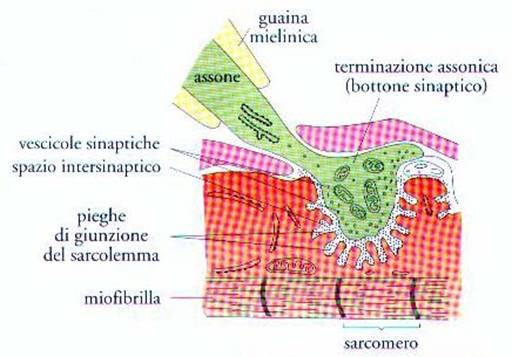
Fig. 8
Propagazione dell’impulso nervoso
Si potrà sottolineare a questo punto, l’importanza
che il sistema muscolare riveste nelle relazioni dell’organismo con l’ambiente
che lo circonda. I muscoli infatti, sono gli organi destinati a mantenere l’equilibrio
nella stazione eretta, a mantenere la postura di una parte qualsiasi del corpo,
a promuovere gli spostamenti del corpo o di una sua parte, opponendosi agli
effetti della gravità. L’azione muscolare é modulata e coordinata di
continuo a livello del sistema nervoso centrale. Le emissioni motrici si
scaricano dal sistema nervoso centrale sulle fibre muscolari striate tramite i
neuroni motori. Il sistema nervoso centrale diventa così responsabile del “tono
muscolare”, cioè della leggera tensione che i muscoli striati presentano a
riposo e che viene rivelata dalla loro caratteristica consistenza e “trofia”.
La contrazione tonica, consente l’attività posturale dei muscoli che fissa le
articolazioni in posizioni determinate e compone l’attitudine d’insieme; é
la tensione dei muscoli che mantiene correttamente le posizioni relative delle
parti del corpo e si oppone alle modificazioni passive di queste posizioni.
Energia della contrazione
muscolare
Come ultima tappa del nostro percorso, rimane da
chiedersi da dove provenga il “carburante” che i muscoli utilizzano per
trarre l’energia necessaria alla loro contrazione. Abbiamo visto come l’ATP
sia necessario per la contrazione muscolare e svolge diversi compiti. E’
necessario quindi che esso sia sempre presente all’interno della cellula
muscolare, ma la concentrazione di ATP all’interno di questa è piuttosto
bassa, per cui ne occorre un rifornimento continuo, affinché la contrazione
possa protrarsi.
Da quali fonti, attinge il
muscolo per rifornirsi di ATP?
Per sviluppare
questa questione, ci serviremo della proiezione della Fig.
9 in
cui è riportato lo schema della produzione di ATP nella fibre muscolari.
Cominciamo con
il dire che la prima riserva, a cui il muscolo attinge per ricostruire
immediatamente ATP, è costituita dalle molecole di fosfocreatina (CP) contenute
nel muscolo stesso. La fosfocreatina è legata all’ATP e all’ADP dalla
seguente reazione:
CP
+ ADP = C + ATP
In cui C indica
la creatina e il segno = equivale ad una doppia freccia.
Dalla
conoscenza delle reazioni chimiche, sappiamo che questa relazione indica che se
il muscolo consuma ATP, allora la reazione si sposta verso destra e le riserve
di CP vengono usate per la produzione di ATP. Se invece, in fase di recupero, il
metabolismo produce ATP… allora la reazione si sposta a sinistra e vengono
reintegrate le riserve di CP. Con questo processo, che tra l’altro avviene in
assenza di ossigeno (anaerobiosi), il muscolo è in grado di sopperire al suo
fabbisogno energetico per i primi secondi di contrazione, ma se questa si
protrae, occorre che il muscolo attinga per il suo fabbisogno ad altre fonti
energetiche, più lente a mettersi in moto, ma più durature.
Esistono
allora, altre due vie di produzione energetica, che non sono fondate su semplici
reazioni ma su “catene di reazioni”, queste sono:
-
la glicolisi
-
la
fosforilazione ossidativa
Non
analizzeremo queste reazioni nel dettaglio in questa sede, anche perché sono
state già trattate in precedenza, ma analizzeremo le caratteristiche salienti
di ognuna, mettendone in evidenza gli aspetti positivi o negativi, e i motivi
per quali viene innescata l’una o l’altra serie di reazioni, a seconda delle
necessità.
La glicolisi
è più rapida, in quanto vi intervengono una serie più breve di reazioni.
Avviene in assenza di ossigeno ma è poco efficiente, in quanto da una molecola
di glucosio, si ottengono solo due molecole di ATP ed inoltre, ha come
catabolita l’acido lattico, il cui accumulo può causare l’abbassamento del
pH nel muscolo, che tende a bloccare la reazione di “distacco” tra i
filamenti actomiosinici.
La fosforilazione
ossidativa è molto efficiente, dato che da una sola molecola di glucosio,
produce 38 molecole di ATP (in realtà sono 36 + 2, cioè andrebbero tolte
quelle che si sono prodotte nella fase glicolitica) però per avvenire, ha
bisogno dell’apporto di ossigeno dal sangue e poi è molto lenta, in quanto vi
è implicata una serie molto più lunga di reazioni.
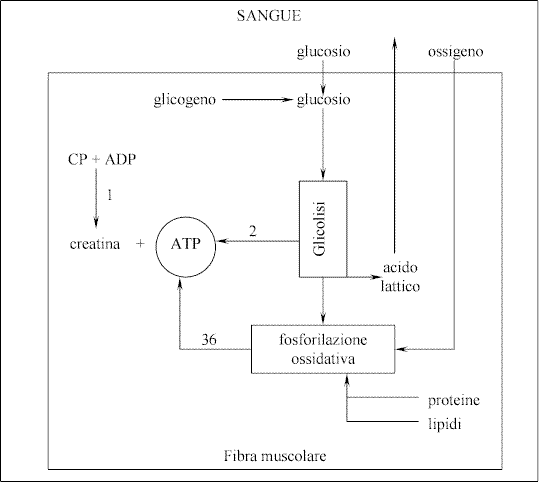
Fig. 9
Vie della produzione di ATP all’interno delle fibre muscolari
Quindi, per un’attività
muscolare lenta e moderata, con un apporto di ossigeno sufficiente, è la
fosforilazione ossidativa a soddisfare il fabbisogno di ATP. Ma per un’attività
sostenuta, diventa importante l’apporto di ATP da parte della glicolisi, che
è rapida e non richiede ossigeno. Il glucosio utilizzato nelle due serie di
reazioni, come schematizzato in Fig. 9, può provenire o dall’apporto del
sangue o dalle riserve di glicogeno accumulate nel muscolo stesso. La
fosforilazione ossidativa, può usare anche proteine e lipidi, la cui
demolizione però, richiede un notevole consumo di ossigeno. Se durante le fasi
di contrazione, il muscolo esaurisce le sue riserve di ATP e CP senza che il
metabolismo sia in grado di reintegrarle, o se l’accumulo di acido lattico
supera una certa soglia… allora si entra nella cosiddetta “fase di fatica”,
in cui il muscolo non è più in grado di contrarsi. La fase successiva, in cui
il muscolo torna a riposo, reintegra le sue riserve e smaltisce l’acido
lattico, viene detta “fase di ristoro”.
Vascolarizzazione e innervazione dei muscoli
Come
collegamento all’unità didattica sulla circolazione sanguigna, anche se non
ancora trattata, si potrà già dire che i muscoli scheletrici sono molto ricchi
di vasi sanguigni. Aree di penetrazione vascolare, uniche o multiple, sono
distribuite generalmente sulla faccia profonda dei muscoli; di qui le arterie
muscolari si irradiano nell’ambiente perifascicolare e interfascicolare, per
risolversi poi, in fitte reti di capillari a maglie allungate, intorno alle
singole fibre muscolari. Le vene hanno un decorso corrispondente a quello delle
arterie. Le fibre nervose destinate ad un muscolo, vi giungono insieme con i
vasi; alcune di esse, dette viscerali, sono appunto destinate all’innervazione
di tali vasi. Le fibre nervose provengono da un nervo spinale, o da più di un
nervo spinale, oppure da nervi encefalici (per i muscoli della testa e per
qualche muscolo del collo).
Bibliografia
-
Alberghina
L., Tonini F. (2002) - “Biologia. Volume b: Dall’evoluzione biologica all’evoluzione
culturale dell’uomo”. Arnoldo Mondadori Scuola, Milano. pp. 622 - 628.
-
Cremaschi
D. (1991) - “Fisiologia generale, principi”. Edi - Ermes, Milano. pp.621 -
642.
-
Curtis
H., N. S. Barnes (2003) - “Invito alla biologia”. Zanichelli, Bologna. pp.337
- 346 e pp. 477 - 481.
-
Voet
D. Voet J.G. (1993) - “Biochimica”. Zanichelli, Bologna. pp. 1290 - 1302.
|
